Để nói về 20 axit amin tạo nên cấu trúc protein và những axit amin đã biến đổi, cần phải mô tả ít nhất 12 con đường trao đổi chất chuyên biệt.
Nhưng tại sao các tế bào sử dụng rất nhiều con đường trao đổi chất đòi hỏi năng lượng (ví dụ để tái tạo các vị trí xúc tác của các enzym), mỗi con đường có một hệ thống enzym, để dị hóa các axit amin? Từ hầu hết tất cả các axit amin, người ta có thể thu được, thông qua các con đường chuyên biệt, các chất chuyển hóa mà một phần nhỏ được sử dụng để sản xuất năng lượng (ví dụ, thông qua quá trình tạo glucone và con đường của các thể xeton) nhưng trên hết, dẫn đến sự hình thành của phức các phân tử, với số lượng nguyên tử cacbon cao (ví dụ từ phenylalanin và tyrosine, các hormone được sản xuất trong tuyến thượng thận chuyên dùng cho mục đích này); nếu một mặt việc sản xuất năng lượng từ các axit amin sẽ đơn giản, mặt khác sẽ rất phức tạp khi xây dựng các phân tử phức tạp bắt đầu từ các phân tử nhỏ: quá trình dị hóa các axit amin cho phép chúng khai thác bộ xương của mình để có được những loài lớn hơn.
Hai hoặc ba héc-ta axit amin bị phân huỷ hàng ngày bởi một người khoẻ mạnh: 60-100 g trong số đó bắt nguồn từ các protein được đưa vào chế độ ăn uống nhưng hơn 2 héc-ta thu được từ sự luân chuyển bình thường của các protein là một phần không thể thiếu của sinh vật ( các axit amin của các protein này, bị phá hủy bởi quá trình oxy hóa khử, được thay thế bằng các axit khác và bị dị hóa).
Các axit amin đóng góp năng lượng về mặt ATP: sau khi loại bỏ nhóm α-amino, bộ xương cacbon còn lại của các axit amin, sau sự biến đổi thích hợp, có thể đi vào chu trình krebs. Hơn nữa, khi nguồn cung cấp chất dinh dưỡng bị thiếu và số lượng glucose giảm, quá trình tạo glucone được kích hoạt: các axit amin gluconeogenetic được cho là những axit mà sau khi sửa đổi thích hợp, có thể được đưa vào gluconeogenesis; axit amin gluconeogenetic là những axit có thể được chuyển đổi thành pyruvate hoặc trong fumarate (fumarate có thể được chuyển thành malate rời khỏi ty thể và trong tế bào chất, được chuyển thành oxaloacetate từ đó có thể thu được phosphoenol pyruvate) giấm-acetate.
Một khía cạnh vừa được mô tả là một khía cạnh rất quan trọng vì axit amin có thể khắc phục tình trạng thiếu đường trong trường hợp nhịn ăn ngay lập tức; Nếu nhịn ăn kéo dài, sau hai ngày chuyển hóa lipid sẽ bị can thiệp (vì các cấu trúc protein không thể bị tấn công quá nhiều) trong giai đoạn này, vì quá trình tạo glucone rất hạn chế, các axit béo được chuyển hóa thành acetyl coenzyme A và các thể xeton. Từ việc nhịn ăn hơn nữa, não cũng thích nghi để sử dụng các thể xeton.
Sự chuyển nhóm α-amino từ axit amin xảy ra thông qua phản ứng chuyển hóa; trên thực tế, các enzym xúc tác phản ứng này, transaminase (hoặc amino transferase). Các enzym này sử dụng đồng yếu tố enzym gọi là pyridoxal phosphat, chất này can thiệp vào nhóm aldehyde của nó. Pyridoxal phosphat là sản phẩm của quá trình phosphoryl hóa pyridoxine, một loại vitamin (B6) được tìm thấy chủ yếu trong rau.
Transaminase có các đặc tính sau:
Độ đặc hiệu cao đối với một cặp α ketoglutarate-glutamate;
Chúng được đặt theo tên của cặp vợ chồng thứ hai.
Enzyme transaminase luôn liên quan đến cặp α ketoglutarate-glutamate và được phân biệt theo cặp thứ hai tham gia.
Ví dụ:
L "aspartate transaminase tức là GOT (Glutamate-Ossal acetate Transaminase): enzyme chuyển nhóm α-amino từ aspartate thành α-ketoglutarate, thu được oxaloacetate và glutamate.
L "alanin transaminase tức là GTP (Glutamate-Pyruvate Transaminase): enzyme chuyển nhóm α-amino từ "alanin thành" α-ketoglutarate, thu được pyruvate và glutamate.
Các transaminase khác nhau sử dụng α-ketoglurat làm chất nhận nhóm amin của axit amin và chuyển nó thành glutamat; trong khi, các axit amin được tạo thành được sử dụng trong quá trình hình thành thể xeton.
Loại phản ứng này có thể xảy ra theo cả hai hướng vì chúng phá vỡ và hình thành các liên kết có cùng năng lượng.
Các transaminase đều có trong tế bào chất và trong ty thể (chúng hầu hết hoạt động trong tế bào chất) và khác nhau về điểm đẳng điện của chúng.
Các transaminase cũng có thể khử cacboxylat axit amin.
Sẽ phải có một cách để chuyển glutamate trở lại α-ketoglutarate: điều này được thực hiện bằng cách khử amin.
Ở đó glutamate dehydrogenase nó là một enzym có khả năng biến đổi glutamat thành α-ketoglutarate và do đó, chuyển đổi các nhóm amin của axit amin được tìm thấy ở dạng glutamat thành amoniac. Những gì diễn ra là một quá trình oxy hóa khử đi qua α-amino glutarate trung gian: amoniac và α-ketoglutarate được giải phóng và quay trở lại vòng tuần hoàn.
Sau đó, việc loại bỏ các nhóm amin của các axit amin chuyển qua các transaminase (khác nhau tùy theo chất nền) và glutamate dehydrogenase, quyết định sự hình thành amoniac.
Có hai loại glutamate dehydrogenase: tế bào chất và ty thể; đồng yếu tố, cũng là cosubstrate của enzym này là NAD (P) +: glutamate dehydrogenase sử dụng NAD + hoặc NADP + làm chất nhận giảm công suất. Dạng tế bào chất thích, mặc dù không phải chỉ riêng, NADP + trong khi dạng ty thể thích NAD +. Hình thức ti thể có mục đích loại bỏ các nhóm amin: nó dẫn đến sự hình thành amoniac (là cơ chất cho một enzym chuyên biệt trong ti thể) và NADH (được gửi đến chuỗi hô hấp). Dạng tế bào chất hoạt động theo hướng ngược lại, nó sử dụng amoniac và α-ketoglutarate để tạo ra glutamate (có đích sinh tổng hợp): phản ứng này là sinh tổng hợp giảm và đồng yếu tố được sử dụng là NADPH.
Glutamate dehydrogenase hoạt động khi cần loại bỏ các nhóm amin của axit amin như amoniac (qua nước tiểu) hoặc khi bộ xương của axit amin cần thiết để sản xuất năng lượng: do đó, enzyme này sẽ có vai trò là bộ điều biến tiêu cực đối với các hệ thống là dấu hiệu sẵn có năng lượng tốt (ATP, GTP và NAD (P) H) và là bộ điều biến tích cực, các hệ thống chỉ ra nhu cầu năng lượng (AMP, ADP, GDP, NAD (P) +, axit amin và hormone tuyến giáp).
Axit amin (chủ yếu là leucine) là chất điều biến tích cực của glutamate dehydrogenase: nếu axit amin có trong tế bào chất, chúng có thể được sử dụng để tổng hợp protein, hoặc chúng phải được thải bỏ vì chúng không thể tích lũy được (điều này giải thích tại sao axit amin là chất điều biến tích cực ).
Xử lý amoniac: chu trình urê
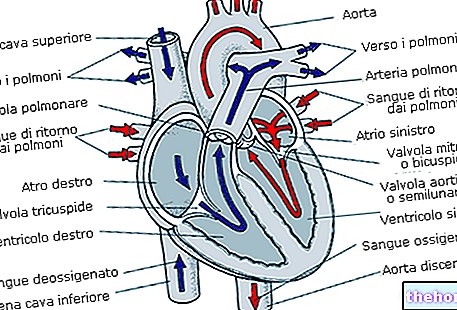
Cá thải amoniac bằng cách đưa nó vào nước qua mang; chim chuyển hóa nó thành axit uric (là một sản phẩm ngưng tụ) và loại bỏ nó bằng phân. Hãy xem điều gì xảy ra ở người: chúng tôi đã nói rằng glutamate dehydrogenase chuyển đổi glutamate thành α- ketoglutarate và amoniac nhưng chúng tôi chưa nói rằng điều này chỉ xảy ra trong ty thể của gan.
Vai trò cơ bản của việc thải amoniac, thông qua chu trình urê, được thực hiện bởi các transaminase của ty thể.

Carbon dioxide, ở dạng ion bicarbonat (HCO3-), được kích hoạt bởi đồng yếu tố biotin tạo thành carboxy biotin phản ứng với amoniac để tạo ra axit cacbamic; phản ứng tiếp theo sử dụng ATP để chuyển một photphat vào axit cacbamic tạo thành cacbamyl photphat và ADP (sự chuyển đổi ATP thành ADP là động lực để thu được carboxybiotine). Giai đoạn này được xúc tác bởi carbamyl phosphate synthetase và xảy ra trong ty thể. Carbamyl phosphate và ornithine là chất nền cho enzym ornithine trans carbamylase chuyển đổi chúng thành citrulline; phản ứng này xảy ra trong ty thể (tế bào gan). Citrulline được tạo ra rời khỏi ty thể và trong tế bào chất, đi dưới "hành động của"arginine succinate synthetase: có sự hợp nhất giữa khung xương cacbon của citrulline và của aspartate thông qua một cuộc tấn công nucleophin và sau đó loại bỏ nước. Enzyme arginine succinate synthetase đòi hỏi một phân tử ATP nên có một sự kết hợp năng lượng: sự thủy phân ATP thành AMP và pyrophosphate (sau đó được chuyển thành hai phân tử orthophosphate) xảy ra bằng cách trục xuất một phân tử nước ra khỏi chất nền và không phải do tác dụng của nước trong môi trường.
"Enzyme tiếp theo là"arginine succinase: enzyme này có thể phân tách arginine succinate thành arginine và fumarate trong tế bào chất.
Chu trình urê được hoàn thành bởi enzym arginase: thu được urê và ornithin; urê được thận thải bỏ (nước tiểu) trong khi ornithine trở lại ty thể và tiếp tục chu trình.
Chu trình urê chịu sự điều biến gián tiếp bởi arginine: sự tích tụ arginine cho thấy rằng chu trình urê phải được tăng tốc; sự điều hòa của arginine là gián tiếp vì arginine điều hòa tích cực enzym acetyl glutamate synthetase. Chất thứ hai có thể chuyển nhóm acetyl trên nitơ của glutamate: N-acetyl glutamate được hình thành là chất điều biến trực tiếp của enzyme carbamyl-phospho synthetase.
Arginine tích lũy như một chất chuyển hóa của chu trình urê nếu việc sản xuất carbamyl phosphate không đủ để thải bỏ ornithine.
Urê chỉ được sản xuất ở gan nhưng có những vị trí khác diễn ra các phản ứng ban đầu.
Bộ não và cơ bắp sử dụng các chiến lược đặc biệt để loại bỏ các nhóm amin. Bộ não sử dụng một phương pháp rất hiệu quả trong đó sử dụng một loại enzyme glutamine synthetase và một loại enzyme glutamase: chất đầu tiên có trong tế bào thần kinh, còn chất thứ hai được tìm thấy trong gan. Cơ chế này rất hiệu quả vì hai lý do:
Hai nhóm amin được vận chuyển từ não đến gan trong một phương tiện duy nhất;
Glutamine ít độc hơn nhiều so với glutamate (glutamate cũng thực hiện chuyển giao tế bào thần kinh và không được vượt quá nồng độ sinh lý).
Ở cá, một cơ chế tương tự đưa nhóm axit amin đến mang.
Từ cơ (xương và tim), các nhóm amin đến gan thông qua chu trình glucose-alanin; enzyme liên quan là glutamine-pyruvate transaminase: nó cho phép chuyển vị các nhóm amin (ở dạng glutamate), chuyển hóa pyruvate thành alanin và đồng thời, glutamate thành α-ketoglutarate trong cơ và xúc tác quá trình ngược lại ở gan.
Các transaminase với các nhiệm vụ hoặc vị trí khác nhau cũng có sự khác biệt về cấu trúc và có thể xác định được bằng phương pháp điện di (chúng có các điểm đẳng điện khác nhau).
Sự hiện diện của transaminase trong máu có thể là một triệu chứng của tổn thương gan hoặc tim (tức là tổn thương mô đối với tế bào gan hoặc tim); các transaminase có nồng độ rất cao ở cả gan và tim: thông qua điện di, có thể xác định xem tổn thương đã xảy ra ở tế bào gan hay tim hay chưa.