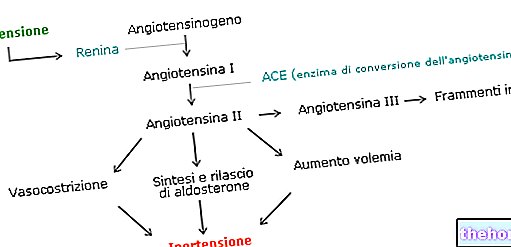
" phần đầu tiên
Enzyme xúc tác giai đoạn thứ tư của chu trình krebs là α-keto glutarate dehydrogenase; enzym này là một phức hợp enzym rất giống với pyruvate dehydrogenase. Cả hai đều bao gồm 48-60 protein, trong đó ba hoạt động enzym khác nhau được công nhận và cũng có các đồng yếu tố enzym giống nhau; là các enzym rất giống nhau vì chúng hoạt động trên các chất nền giống nhau: cả pyruvate và l "α-keto glutarate, là α- axit xeto. Cơ chế hoạt động của hai phức hợp enzym là như nhau.
Sự tấn công của thiamine pyrophosphate vào cacbonyl (C = O) của "α-ketoglutarate, dẫn đến quá trình khử cacboxyl của nó và dẫn xuất cacboxyhydroxy propyl được hình thành. Với sự chuyển tiếp sau đó sang lipoamit, quá trình oxy hóa khử bên trong sẽ diễn ra, từ đó thu được dẫn xuất lipoamit cacboxy hoặc succinyl lipoamit.
Sau đó, lipoamit succinyl phản ứng với coenzym A để tạo ra succinyl coenzym A (tiếp tục trong chu trình krebs) và lipoamit bị khử được tái oxy hóa bởi FAD: FADH2 được tạo thành được tái oxy hóa bởi NAD + và thu được NADH. Do đó, trong giai đoạn này, quá trình loại bỏ lần thứ hai carbon khỏi bộ xương carbon đã diễn ra, dưới dạng carbon dioxide.
Một nhóm acyl liên kết với coenzyme A ở dạng hoạt hóa, tức là nó có hàm lượng năng lượng cao: do đó có thể khai thác năng lượng của succinyl coenzyme A.

Trong giai đoạn thứ năm của chu trình krebs, succinyl coenzyme A chịu tác dụng của succinyl thiokinase; hai giả thuyết đã được đưa ra về phương thức hoạt động của nó: chúng tôi sẽ chỉ mô tả một trong hai giả thuyết vì nó được công nhận nhiều nhất. Theo giả thuyết này, succinyl coenzyme A bị tấn công bởi nitơ của histidine (Hys) của enzyme: coenzyme A được giải phóng và một sản phẩm bổ sung có nguồn gốc từ histidine được hình thành như một chất trung gian, đó là succinyl-enzyme (hoặc succinyl-Hys ); một orthophosphate tác động lên chất trung gian này, dẫn đến giải phóng succinate và hình thành phosphoenzyme. Phosphoenzyme, bị tấn công bởi guanosine diphosphate (GDP), tạo ra guasnosine triphosphate (GTP) và enzyme được giải phóng. Theo quan điểm năng lượng GTP = ATP: liên kết cung cấp năng lượng là như nhau ở cả hai loài (đó là liên kết anhydrit giữa photphoryl Β và photphoryl γ). Trong một số trường hợp, GTP được sử dụng làm nguyên liệu có hàm lượng năng lượng cao, nhưng thông thường, GTP được chuyển hóa thành ATP nhờ tác động của enzym nucleoside diphospho kinase (NDPK); là một loại enzym được tìm thấy trong tế bào và xúc tác phản ứng sau:
N1TP + N2DP → N1DP + N2TP
NiTP ® Nucleoside Triphosphate chung
NiDP ® nucleoside diphosphate chung
Đó là một phản ứng thuận nghịch; trong trường hợp của chúng tôi, nó xảy ra:
GTP + ADP → GDP + ATP
do đó nó có thể tiến hành sang phải hoặc sang trái ngay cả khi có sự thay đổi nhỏ về nồng độ của thuốc thử.
Nếu chu trình krebs diễn ra với tốc độ dẫn đến sản xuất ATP cao hơn nhu cầu năng lượng, thì ADP sẽ khan hiếm trong khi có rất nhiều ATP: phản ứng được xúc tác bởi nucleoside diphospho kinase, khi đó, hướng sang trái (GTP tích lũy nếu nucleoside diphospho kinase không có đủ chất nền, tức là ADP). GTP do đó là một tín hiệu về sự sẵn có năng lượng và do đó làm chậm chu kỳ krebs.
Giai đoạn thứ sáu của chu kỳ krebs dẫn đến sự hình thành của fumarate bằng hành động của succinate dehydrogenase; enzym này tạo ra một phản ứng đặc hiệu nổi khi trans không bão hòa (nó là một anken) luôn được hình thành, tức là fumarate (trong khi đồng phân cis là maleat). Succinate dehydrogenase được tìm thấy trên màng trong ty thể, trong khi tất cả các enzym khác của chu trình krebs nằm rải rác khắp ty thể.
Succinate dehydrogenase có FAD như một đồng yếu tố; nó bị ức chế bởi oxaloacetate (ức chế hồi lưu) trong khi nó có succinate và fumarate là chất điều biến tích cực (chất kích hoạt). chất kích hoạt của nó. Hãy thử tìm hiểu lý do tại sao, bằng cách chuyển sang giai đoạn cuối cùng của chu kỳ krebs. Giai đoạn cuối cùng của chu trình krebs đòi hỏi năng lượng vì vậy khả năng duy nhất để lấy oxaloacetate từ bệnh nhân là nồng độ của bệnh nhân rất cao: malate là một trong những chất chuyển hóa có nồng độ cao nhất trong tế bào. Phản ứng chuyển malate thành oxaloacetate cũng được ưa chuộng bởi thực tế là nồng độ của oxaloacetate được giữ ở mức thấp do hoạt động của citrate synthase. Khi đó, phản ứng được xúc tác bởi succinate dehydrogenase là một phản ứng tự ăn và đây là cách duy nhất để diễn ra quá trình chuyển hóa malate thành oxaloacetate.
Nồng độ của malat trong ty thể phải tương thích với nồng độ của malat trong tế bào chất: chỉ khi nồng độ của malat trong ty thể cao đến mức đảm bảo chuyển hóa malat thành oxaloacetat (trong chu trình krebs) thì malat cũng có thể được sử dụng trong các cách khác (là tế bào chất): trong tế bào chất, malate có thể được chuyển đổi thành oxaloacetate từ đó có thể thu được aspartate bằng tác dụng của GOT (nó là một transaminase) hoặc glucose thông qua quá trình gluconeogenesis.
Chúng ta quay trở lại giai đoạn thứ bảy của chu trình krebs, nó được xúc tác bởi enzyme fumarasi: nước được thêm vào một cách cụ thể để tạo ra L-malate.
Trong giai đoạn cuối của chu trình Krebs, mà chúng ta đã nói, hành động của malate dehydrogenase. Enzyme này sử dụng một phân tử NAD + cho hoạt động xúc tác của nó.
Do đó, chúng tôi đã kết luận mô tả về các giai đoạn khác nhau của chu kỳ krebs.
Chu kỳ krebs hoàn toàn có thể đảo ngược.
Để tăng tốc độ của chu trình krebs, có thể tăng nồng độ của các chất chuyển hóa có trong chu trình đó; một trong những chiến lược để tăng tốc độ của chu trình krebs bao gồm chuyển đổi một phần của pyruvate đi vào ty thể thành oxaloacetate (do tác dụng của pyruvate carboxylase) và không biến đổi tất cả thành acetyl coenzyme A: do đó làm tăng nồng độ của oxaloacetate. là một chất chuyển hóa của chu trình krebs và do đó, làm tăng tốc độ của cả chu kỳ.
Trong chu trình krebs, ba NAD + được chuyển thành ba NADH và một FAD thành FADH2 và hơn nữa, GTP thu được: bằng cách phân luồng năng lượng khử thu được từ chu trình krebs, ATP được sản xuất thêm; Trong chuỗi hô hấp, năng lượng khử được chuyển từ NADH và FADH2 thành oxy: sự chuyển này là do một loạt các enzym nằm trên màng ty thể hoạt động dẫn đến sản xuất ATP.
Các quá trình của chuỗi hô hấp là các quá trình gắng sức và năng lượng được giải phóng được sử dụng để sản xuất ATP; mục đích của tế bào là khai thác các quá trình xuất tiết để làm cho quá trình tổng hợp ATP diễn ra. Cứ mỗi phân tử NADH đi vào chuỗi hô hấp thì thu được 2,5 phân tử ATP và cứ FADH2 thì thu được 1,5 phân tử ATP; Sự đa dạng này là do FADH2 đi vào chuỗi hô hấp ở mức độ thấp hơn NADH.
Với khả năng khử của quá trình chuyển hóa hiếu khí, thu được 30-32 ATP (219-233 kcal / mol) với hiệu suất khoảng 33% (hiệu suất của quá trình chuyển hóa kỵ khí khoảng 2%).
.jpg)